Depression is a complex multifactorial neuropsychiatric condition which is associated with a high burden of disease [1-3]. Depression disturbs mood and cognition resulting in poor functioning and a significant detriment to the quality of life [1, 3-5]. It is one of the most prevalent disorders affecting more than 300 million individuals globally [6]. Five per cent of the Australian population suffer from depression each year [7]. Depression is characterised by a high recurrence rate; 40% of the treated patients relapse within 12 months [1]. Moreover, 20% to 30% of patients receiving a treatment for depression, achieve partial remission [1]. These patients continue to experience partial negative symptoms and suffer from daily life limitations. Depressive disorders are heterogenous and include a wide variety of biological and psychopathological endophenotypes [8]. Psychopathological endophenotypes include depressed mood, anhedonia, impaired learning and memory, appetite change, diurnal variation and stress [9]. Biological endophenotypes include REM sleep abnormalities, catecholamine depletion, downregulation of serotonin (5-HT) receptors, tryptophan (L-Trp) depletion and high cortisol levels [9]. The current diagnosis of depressive disorders and monitoring treatment success involve the use of depression assessment scales [9]. These scales are based on subjective markers; patients’ symptoms. Hence, the current diagnosis and treatment of depression can be affected by clinicians’ bias to a higher extent in the absence of accurate biomarkers guiding the diagnosis and treatment of depression.
To date, there are no acknowledged objective markers employed to monitor treatment response of antidepressants in practice. The delayed response of antidepressants and the lack of accurate markers guiding the treatment therapy hinders physicians’ clinical decisions during the initial weeks of treatment. Having objective biological markers to study depressive symptoms can improve the initial treatment of depression by modelling patients’ disease state and their responses to antidepressant interventions.
Initially, depression was linked to a relative deficiency of monoamine neurotransmitters in the synaptic cleft such as 5-HT and noradrenaline (NA) [3, 5]. This is known as the monoamine theory of depression [3, 14]. 5-HT levels were used as a marker to guide the efficacy of treatment for depression. Soon after, researchers discovered the limitations of the monoamine hypothesis. Although, antidepressants medications correct the levels of neurotransmitters at the synapse within a few hours after the first dose, a period of four to six weeks is needed to achieve the full antidepressant effects in improving patients’ mood as outlined in Figure 1 [15]. Additionally, the current antidepressants medications have a success rate of approximately 60% [15]. This highlights a limitation in this theory and suggests the involvement of alternative mechanisms to ameliorate mood other than the direct effect of antidepressants on the level of neurotransmitters at the synapse.
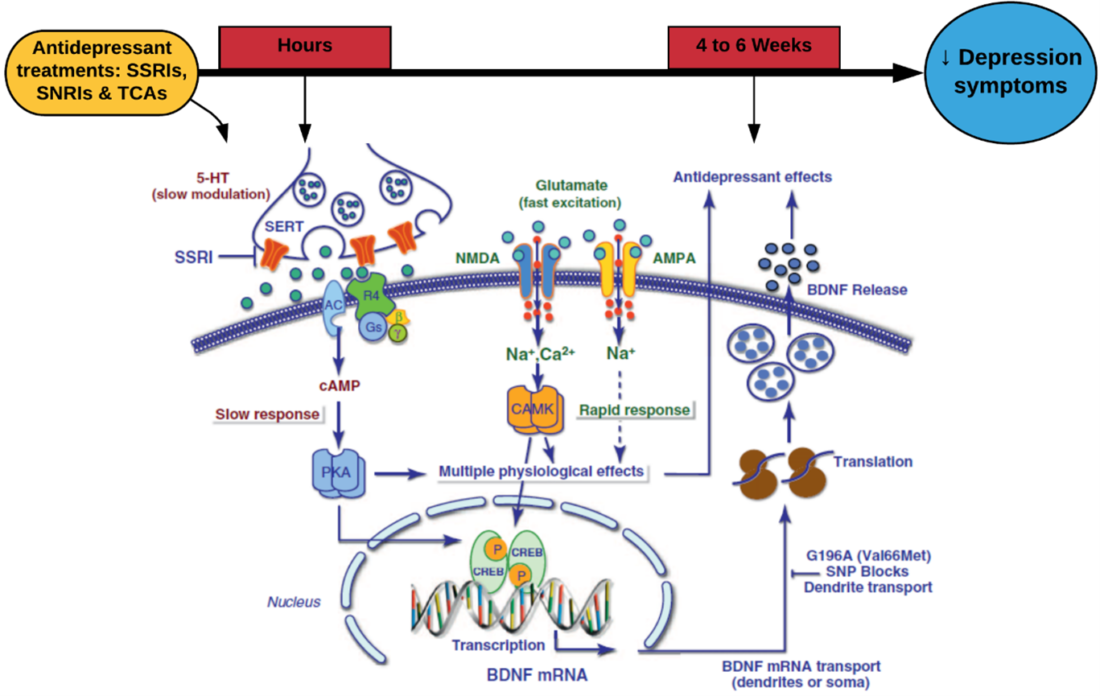
Figure 1 – Overview of mechanisms involved in depression and antidepressants mechanism of actions
Many studies identified relevant biological markers that are associated with depression. Recent evidence suggests a correlation between depression and neuroinflammation causing a disruption to L-tryptophan (Trp) and kynurenine pathway (KP) along with the neopterin to biopterin (N/B) ratio [10-12]. Several studies focused on the Trp depletion and upregulation of the KP in certain endophenotypes of depression whereas limited studies have investigated the potential of using neopterin to biopterin (N/B) ratio as a marker to monitor antidepressants’ treatment response. Evidence suggests that a rise in N/B ratio in depressed patients correlates with the severity depressive symptoms [12].
Overall, depression remains on of the most prevalent conditions with a high disease burden. Current depression treatments have a 60% success rate require four to six weeks to show their full effect. Understanding the different mechanisms that may be involved in the pathophysiology of depression, will assist identifying novel, more effective treatments and relevant biomarkers that can monitor treatment response.
Mina Malek, BBiomedSc, MPharm (Senior Scientific Advisor at My Medical Department)
References:
1. Peter M. Ellis IH, Smith. aDAR. Australian and New Zealand clinical practice guidelines for the treatment of depression Royal Australian and New Zealand College of Psychiatrists Clinical Practice Guidelines Team for Depression. Australian and New Zealand Journal of Psychiatry. 2004;38:389-407.
2. Kaster MP, Moretti M, Cunha MP, Rodrigues ALS. Novel approaches for the management of depressive disorders. European Journal of Pharmacology. 2016;771:236-40.
3. Sahin C, Dursun S, Cetin M, Aricioglu F. The Neuroinflammation Perspective of Depression: Reuniting the Outstanding Mechanisms of the Pathophysiology. 2016:196.
4. Duman RS, Voleti B. Signaling pathways underlying the pathophysiology and treatment of depression: novel mechanisms for rapid-acting agents. Trends in Neurosciences. 2012;35:47-56.
5. Maes M, Mihaylova I, Kubera M, Ringel K. Activation of cell-mediated immunity in depression: Association with inflammation, melancholia, clinical staging and the fatigue and somatic symptom cluster of depression. Progress in Neuro-Psychopharmacology and Biological Psychiatry. 2012;36:169-75.
6. Maes M. Depression is an inflammatory disease, but cell-mediated immune activation is the key component of depression. Progress in Neuro-Psychopharmacology and Biological Psychiatry. 2011;35:664-75.
7. Maes M, Ringel K, Kubera M, Berk M, Rybakowski J. Increased autoimmune activity against 5-HT: A key component of depression that is associated with inflammation and activation of cell-mediated immunity, and with severity and staging of depression. Journal of Affective Disorders. 2012;136:386-92.
8. Maes M, Leonard BE, Myint AM, Kubera M, Verkerk R. The new ‘5-HT’ hypothesis of depression: Cell-mediated immune activation induces indoleamine 2,3-dioxygenase, which leads to lower plasma tryptophan and an increased synthesis of detrimental tryptophan catabolites (TRYCATs), both of which contribute to th. Progress in Neuro-Psychopharmacology and Biological Psychiatry. 2011;35:702-21.
9. Gabbay V, Klein RG, Alonso CM, Babb JS, Nishawala M, De Jesus G, et al. Immune system dysregulation in adolescent major depressive disorder. Journal of affective disorders. 2009;115:177-82.
10. Raison CL, Miller AH. Is depression an inflammatory disorder? Current Psychiatry Reports. 2011;13:467-75.
11. Hall S, Arora D, Anoopkumar-Dukie S, Grant GD. Effect of Coffee in Lipopolysaccharide-Induced Indoleamine 2,3-Dioxygenase Activation and Depressive-like Behavior in Mice. Journal of Agricultural and Food Chemistry. 2016;64:8745-54.
12. Sublette ME, Postolache TT. Neuroinflammation and Depression. Psychosomatic Medicine. 2012;74:668-72.
13. Gabbay V, Klein RG, Katz Y, Mendoza S, Guttman LE, Alonso CM, et al. The possible role of the kynurenine pathway in adolescent depression with melancholic features. Journal of Child Psychology and Psychiatry. 2010;51:935-43.
14. Réus GZ, Jansen K, Titus S, Carvalho AF, Gabbay V, Quevedo J. Kynurenine pathway dysfunction in the pathophysiology and treatment of depression: Evidences from animal and human studies. Journal of Psychiatric Research. 2015;68:316-28.
15. Raison CL, Dantzer R, Kelley KW, Lawson MA, Woolwine BJ, Vogt G, et al. CSF concentrations of brain tryptophan and kynurenines during immune stimulation with IFN-α: relationship to CNS immune responses and depression. Molecular Psychiatry. 2010;15:393-403.